400-998-5282
专注多肽 服务科研
400-998-5282
专注多肽 服务科研
甲状旁腺激素(PTH),又称为甲状旁腺激素或甲状旁腺,是由甲状旁腺主细胞分泌的含有84个氨基酸的多肽。它的作用是增加血液中钙的浓度,PTH通过作用于甲状旁腺激素1受体和甲状旁腺激素2受体,增加血液中的钙浓度(1)。 PTH降低肾脏的近端小管磷酸盐的再吸收,这意味着更多的磷酸盐通过尿排出体外。PTH增加从肠道和骨头吸收进入血液的磷酸盐含量(2)。在骨中,从破碎的骨中释放的钙的含量比磷酸盐稍多。在肠内,活性维生素D增加,介导钙和磷的吸收。
促甲状腺激素释放激素(TRH)的定义
促甲状腺激素释放激素(TRH)是一种神经激素,是最简单的下丘脑神经激素。它由谷氨酸-组氨酸-脯氨酸序列中的三个氨基酸组成。TRH的结构简单是有欺骗性的,因为这种激素实际上具有许多功能。它刺激垂体前叶合成和分泌促甲状腺激素【1】。
Thyrotropin-releasing hormone (TRH), a neurohormone, is the simplest of the hypothalamic neuro- hormones. It consists of three amino acids in the sequence glutamic acid–histidine–proline. The structural simplicity of TRH is deceiving because this hormone actually has many functions. It stimulates the synthesis and secretion of thyrotropin by the anterior pituitary 【1】.
促甲状腺激素释放激素(TRH)的相关肽
TRH的发现很快导致了其他小肽的发现,这些小肽调节垂体前叶分泌其热带激素。这种调节通常通过增加或减少刺激来完成。然而,生长激素似乎主要通过增加或减少抑制来调节,尽管已经确定了生长激素释放因子。TRH首先被鉴定的肽物质通常被归类为下丘脑垂体激素【1】。
The discovery of TRH led quickly to the discovery of other small peptides that regulate the secretion by the anterior pituitary of its tropic hormones. Such regulation is usually accomplished by increasing or decreasing stimulation. However, growth hormone appears mainly to be regulated by increasing or decreasing inhibition, though a growth-hormone-releasing factor has been identified. The peptide substances of which TRH was the first to be identified are often grouped as hypothalamic hypophysiotropic hormones 【1】.
促甲状腺激素释放激素(TRH)的发现
1969年,由Guillemin和Schally领导的一个小组在竞争性工作多年后宣布,导致垂体前叶释放促甲状腺激素(促甲状腺激素,TSH)的下丘脑物质是L-焦谷氨酰-L-组氨酸-L-脯氨酸-酰胺(L-pGlu-L-His-L-ProNH2)。这种三肽现在被称为促甲状腺激素释放激素(TRH)【1】。
In 1969, a group led by Guillemin and another by Schally, having worked competitively for many years, announced that the hypothalamic substance that causes the anterior pituitary gland to release thyrotropin (thyroid-stimulating hormone, TSH) is L-pyroglutamyL-L-histidyl-L-proline- amide (L-pGlu-L-His-L-ProNH2). This tripeptide is now called thyrotropin-releasing hormone (TRH) 【1】.
促甲状腺激素释放激素(TRH)的结构特征
使用分子技术证实了两栖动物中TRH合成的翻译后酶促模式。确定青蛙DNA含有478个核苷酸的片段,编码pro-TRH的氨基末端区域,这是一个123个氨基酸的前体,包含TRH祖细胞序列(Gln-His-pro-Gly)的三个拷贝,两侧是成对的二元残基,第四个不完整拷贝缺少C端甘氨酸。后来在大鼠下丘脑中鉴定出哺乳动物促TRH分子为255个氨基酸的蛋白质,其中包含TRH祖细胞氨基酸序列的五个拷贝。人类pro-TRH包含六个副本。TRH的生物合成基本上是一个五步过程,从TRH基因的DNA转录成细胞核内的TRH mRNA开始。转录后,将TRH mRNA翻译成核糖体上的前TRH肽。TRH的翻译后加工始于通过羧基肽酶的作用切除祖肽。然后通过肽基甘氨酸α酰胺化单加氧酶对脯氨酸进行酰胺化,酰胺部分由C端甘氨酸提供。最后,通过谷氨酰胺环化酶完成N-末端谷氨酰胺的环化。由于在正中隆起或脊髓的轴突或末端缺乏TRH祖细胞免疫反应性,TRH的翻译后加工似乎仅限于神经元核周。pro-TRH的翻译后加工还产生了许多其他可能具有行为或生理活性的肽【1】。
A post-translational enzymatic mode of TRH synthesis in amphibians was confirmed using molecular techniques. It was determined that frog DNA contained a segment of 478 nucleotides that coded for the amino-terminal region of pro-TRH, a 123-amino-acid precursor containing three copies of the progenitor sequence of TRH (Gln-His-Pro-Gly) flanked by paired dibasic residues and a fourth incomplete copy lacking the C-terminal glycine. A mammalian pro-TRH molecule was later identified in rat hypothalamus as a 255-amino-acid protein containing five copies of the amino acid sequence of the TRH progenitor. Human pro-TRH contains six copies. The biosynthesis of TRH is essentially a five-step process, beginning with transcription of DNA of the TRH gene to TRH mRNA within the cell nucleus. Transcription is followed by translation of the TRH mRNA to the pro-TRH peptide on the ribosome. The post-translational processing of TRH begins with excision of the progenitor peptides by the action of carboxy peptidases. This is followed by amidation of proline by peptidyl glycine alpha-amidating monoxygenase, the amide moiety being donated by the C-terminal glycine. Finally, cyclization of the N-terminal glutamine by glutaminyl cyclase is accomplished. Post-translational processing of TRH appears to be restricted to the neuronal perikarya because of lack of TRH progenitor immunoreactivity in axons or terminals of the median eminence or spinal cord. The post-translational processing of pro-TRH also gives rise to a number of other peptides that may have behavioral or physiological activity 【1】.
促甲状腺激素释放激素(TRH)的作用机制
TRH受体存在于垂体前叶的促甲状腺激素和乳腺营养细胞以及整个中枢神经系统的神经元上。垂体和中枢TRH受体的结构和功能特性已被详细表征,并显示出在结构,结合特征和信号转导机制上相似。TRH受体属于一类G蛋白偶联受体,具有七个跨膜结构域和一个包含N-糖基化位点的细胞外N端区域。已经使用培养的垂体瘤细胞研究了TRH作用于TSH和PRL分泌的基本细胞机制。这两种激素的双相分泌已被证明涉及G蛋白偶联刺激肌醇磷脂周转。激素分泌的初始阶段被认为是由肌醇三磷酸介导的细胞内储存释放Ca2+引起的;第二个也是更持续的阶段是由于细胞外Ca2+通过由1,2-DG诱导的蛋白激酶C激活的电压敏感通道流入。刺激促甲状腺激素肿瘤细胞上的TRH受体也诱导花生四烯酸代谢物的释放,花生四烯酸代谢物可能充当额外的细胞内信使。除了刺激TSH和PRL释放外,TRH还调节PRL的转录和TSH 的翻译后糖基化【1】。
Receptors for TRH are found on thyrotroph and mammotroph cells of the anterior pituitary and on neurons throughout the CNS. The structural and functional properties of both pituitary and central TRH receptors have been characterized in detail and shown to be similar in structure, binding characteristics, and mechanisms of signal transduction. TRH receptors belong to a class of G-protein-coupled receptors with seven membrane-spanning domains and an extracellular N-terminal region containing N-glycosylation sites. The basic cellular mechanisms of TRH actions on TSH and PRL secretion have been investigated using cultured pituitary tumor cells. The biphasic secretion of these two hormones has been shown to involve G-protein-coupled stimulation of inositol phospholipid turnover. The initial phase of hormone secretion is thought to result from inositol trisphosphate-mediated release of Ca2+ from intracellular stores; the second and more sustained phase results from influx of extracellular Ca2+ via voltage-sensitive channels activated by protein kinase C, which is induced by 1,2-DG. Stimulation of TRH receptors on thyrotroph tumor cells also induces the release of arachidonic acid metabolites that may act as additional intracellular messengers. In addition to stimulating TSH and PRL release, TRH regulates the transcription of PRL and the post-translational glycosylation of TSH【1】.
某些低亲和力TRH类似物如MeTRH,TRH-Gly(pGlu-His-Pro-GlyOH)和Phe2-TRH(pGlu-Phe-ProNH2),NP654(pGlu-His(1-异丙基)-ProNH2),3R-Desaza-TRH((1R)-(3-氧代环戊基)-His-ProNH2)和S-Desaza-TRH((1S)-(3-氧代环戊基)-His-ProNH2)是TRH-R1和TRH-R2比同源配体TRH 更有效的激动剂【2】。
Certain low affinity TRH analogs such as MeTRH, TRH-Gly (pGlu-His-Pro-GlyOH) and Phe2-TRH (pGlu-Phe-ProNH2), NP654 (pGlu-His(1-isopropyl)-ProNH2), 3R-Desaza-TRH ((1R)-(3-oxocyclopentyl)-His-ProNH2), and S-Desaza-TRH ((1S)-(3-oxocyclopentyl)-His-ProNH2) are more efficacious agonists at TRH-R1 and TRH-R2 than the cognate ligand TRH 【2】.
促甲状腺激素释放激素(TRH)的功能
垂体前叶功能:TRH明确的内分泌功能是刺激垂体前叶促甲状腺细胞合成和释放TSH。因此,TRH与甲状腺激素以及下丘脑多巴胺和生长抑素的抑制作用一起,控制垂体TSH的合成和释放。TRH刺激催乳素(PRL)的释放,但它是主要的生理性PRL释放因子的证据是物种特异性的或有争议的。
Anterior Pituitary Function: The unequivocal endocrine function of TRH is to stimulate the synthesis and release of TSH from thyrotroph cells of the anterior pituitary gland. Thus, TRH, in concert with thyroid hormones and the inhibitory influences of dopamine and somatostatin from the hypothalamus, controls pituitary TSH synthesis and release. TRH stimulates prolactin (PRL) release, but evidence that it is a major physiologic PRL-releasing factor is either species-specific or controversial.
中枢作用:TRH的中枢作用是无数的,影响大脑的化学,生理和行为。作为几种不同神经递质的神经调节剂,包括最显着的多巴胺,血清素,乙酰胆碱和阿片类药物,TRH影响许多药物的作用,这些药物本身会影响这些和其他神经递质。
Central Actions: The central actions of TRH are myriad, affecting brain chemistry, physiology, and behavior. As a neuromodulator of several different neurotransmitters, including most prominently dopamine, serotonin, acetylcholine, and the opiates, TRH affects the actions of many drugs that themselves affect these and other neuro-transmitters.
TRH已被证明可以通过海马机制唤醒冬眠动物,并拮抗乙醇和其他中枢神经系统抑制剂产生的镇静,运动障碍和低温。TRH抵消由各种药物和几种内源性神经肽(包括神经降压素,蛙皮素和β-内啡肽)产生的低温或变温。它单独对体温的影响是可变的:TRH在某些物种中产生体温过低,在其他物种中产生体温过高,而在某些物种中它没有影响。类似地,TRH通过激活中脑边缘多巴胺系统来刺激运动活动。三肽还对心血管和呼吸系统产生深刻的刺激,并诱导胃肠动力增加以及胃液的体积和酸度,同时通常抑制食物和水的摄入。TRH的这些胃肠道作用可能在TRH的溃疡形成作用中起作用。
TRH has been shown to arouse hibernating animals, through a hippocampal mechanism, and to antagonize the sedation, motor impairment, and hypothermia produced by ethanol and other CNS depressants. TRH counteracts the hypothemia or poikilothermia produced by various drugs and several endogenous neuropeptides, including neurotensin, bombesin, and betaendorphin. Its effects alone on body temperature are variable: TRH produces hypothermia in some species, hyperthermia in others, and in some it has no effect. Similarly, TRH stimulates locomotor activity by activation of the mesolimbic dopamine system. The tripeptide also produces profound stimulation of the cardiovascular and respiratory systems and induces increases in gastrointestinal motility and the volume and acidity of gastric secretion while often suppressing the intake of both food and water. These gastrointestinal effects of TRH may play a role in the ulcerogenic actions of TRH.
在神经系统疾病中:如上所述,在发现TRH作为下丘脑促垂体激素后不久,发现TRH分布在大脑,脊髓和其他器官系统的许多下丘脑外部位。TRH的神经解剖学位置和神经化学作用表明它可以用作神经和精神疾病的治疗剂【1】。
In Neurological Disorders: As described above, shortly after its discovery as a hypothalamic hypophysiotropic hormone, TRH was found to be distributed in many extrahypothalamic sites in brain, in spinal cord, and in other organ systems. The neuroanatomical location and neurochemical actions of TRH suggested that it could be utilized as a therapeutic agent in neurological and psychiatric disorders 【1】.
促甲状腺激素释放激素(TRH)的相关文献
1. Book: Focus on Basic Neurobiology. Thyrotropin-Releasing Hormone by Mason GA, Garbutt JC, Prange, Jr AJ (2000).
2. Engel S, Neumann S, Kaur N, Monga V, Jain R, Northup JGershengorn MC (2006). Low Affinity Analogs of Thyrotropin-releasing Hormone Are Super-agonists. The Journal of Biological Chemistry, 281(19):13103-13109.
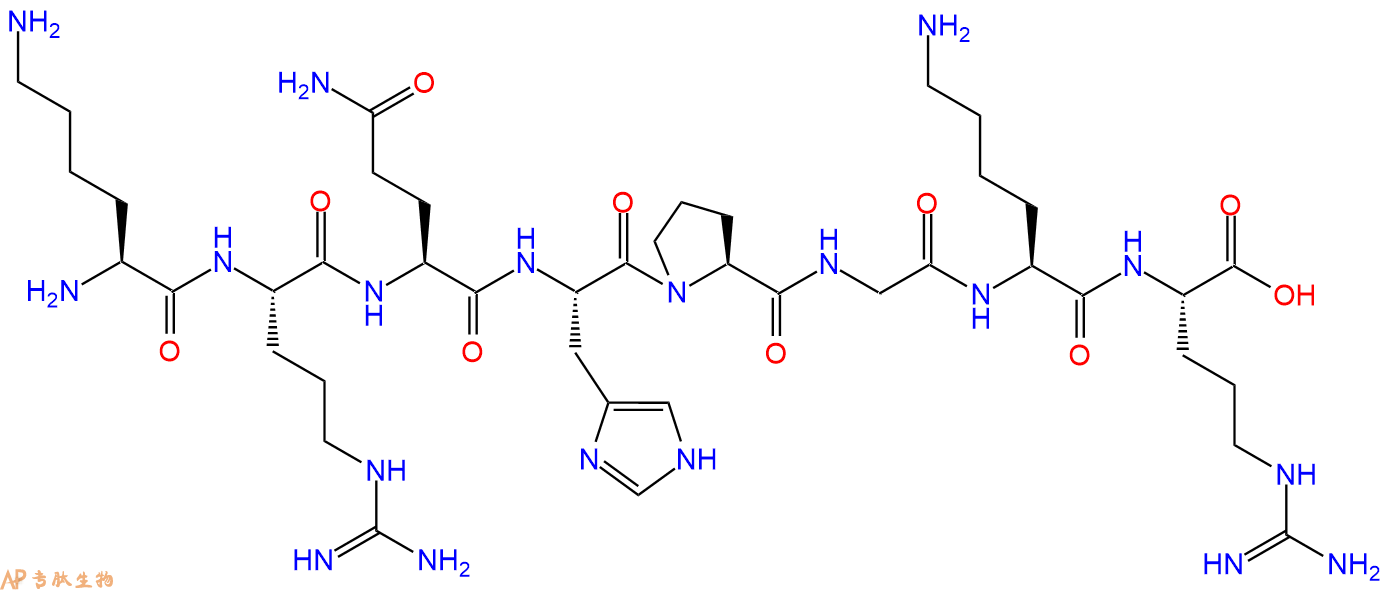
多肽H2N-Lys-Arg-Gln-His-Pro-Gly-Lys-Arg-COOH的合成步骤:
1、合成CTC树脂:称取0.96g CTC Resin(如初始取代度约为0.77mmol/g)和0.89mmol Fmoc-Arg(Pbf)-OH于反应器中,加入适量DCM溶解氨基酸(需要注意,此时CTC树脂体积会增大好几倍,避免DCM溶液过少),再加入2.22mmol DIPEA(Mw:129.1,d:0.740g/ml),反应2-3小时后,可不抽滤溶液,直接加入1ml的HPLC级甲醇,封端半小时。依次用DMF洗涤2次,甲醇洗涤1次,DCM洗涤一次,甲醇洗涤一次,DCM洗涤一次,DMF洗涤2次(这里使用甲醇和DCM交替洗涤,是为了更好地去除其他溶质,有利于后续反应)。得到 Fmoc-Arg(Pbf)-CTC Resin。结构图如下:
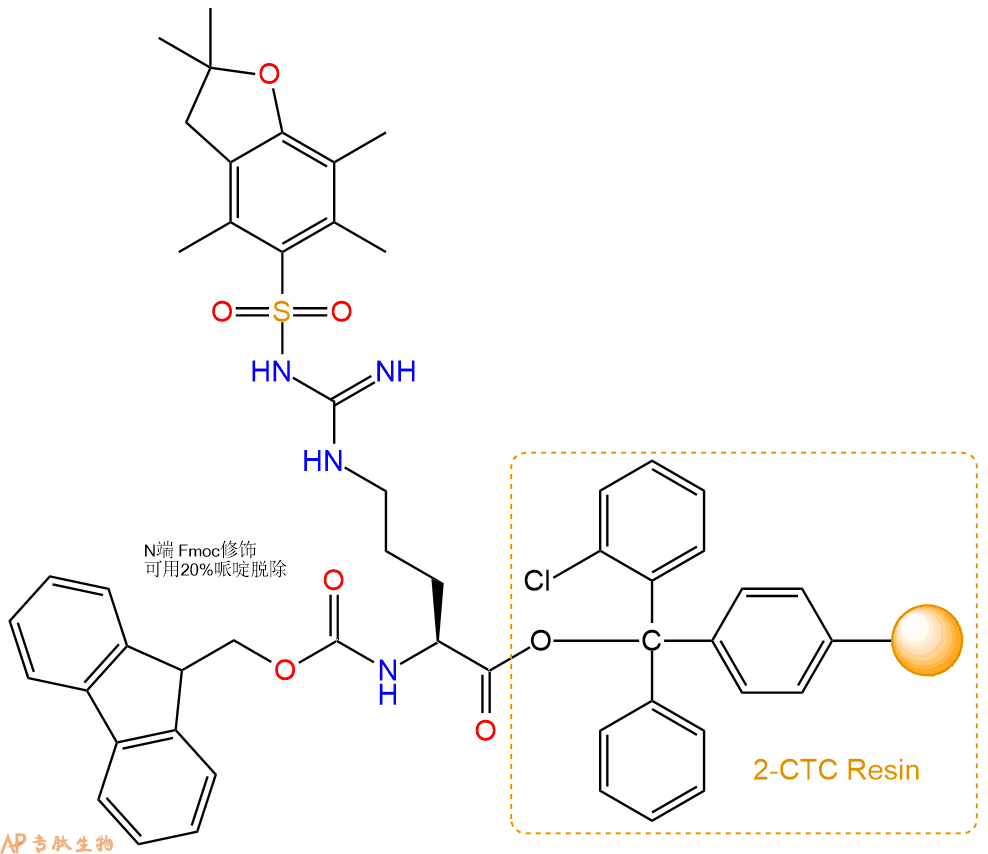
2、脱Fmoc:加3倍树脂体积的20%Pip/DMF溶液,鼓氮气30分钟,然后2倍树脂体积的DMF 洗涤5次。得到 H2N-Arg(Pbf)-CTC Resin 。(此步骤脱除Fmoc基团,茚三酮检测为蓝色,Pip为哌啶)。结构图如下:
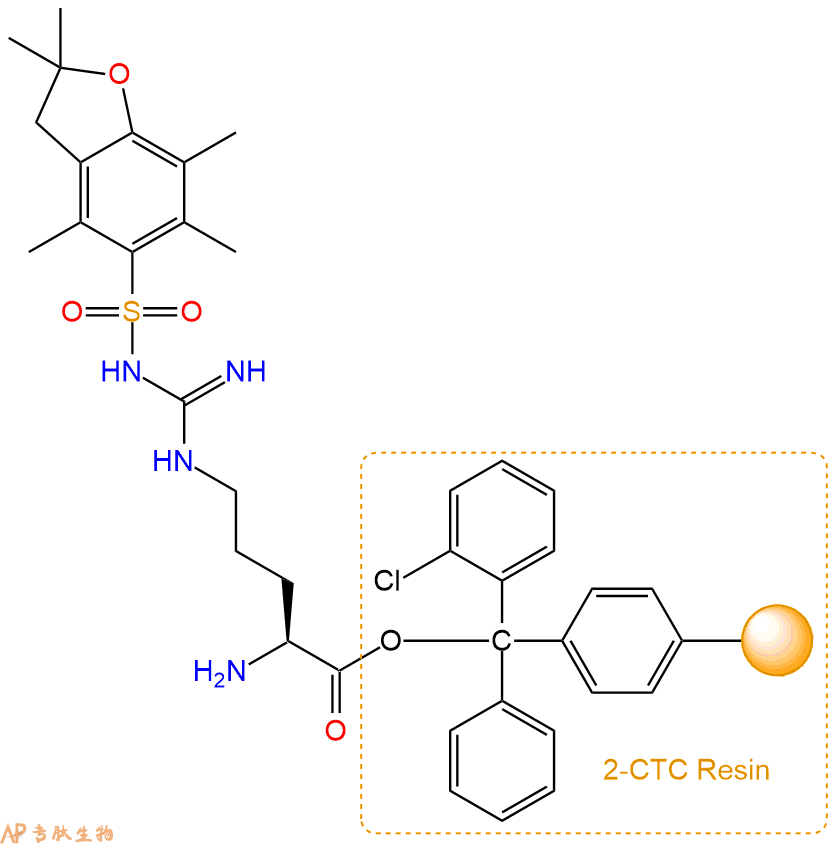
3、缩合:取2.22mmol Fmoc-Lys(Boc)-OH 氨基酸,加入到上述树脂里,加适当DMF溶解氨基酸,再依次加入4.44mmol DIPEA,2.11mmol HBTU。反应30分钟后,取小样洗涤,茚三酮检测为无色。用2倍树脂体积的DMF 洗涤3次树脂。(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。得到Fmoc-Lys(Boc)-Arg(Pbf)-CTC Resin。氨基酸:DIPEA:HBTU:树脂=3:6:2.85:1(摩尔比)。结构图如下:
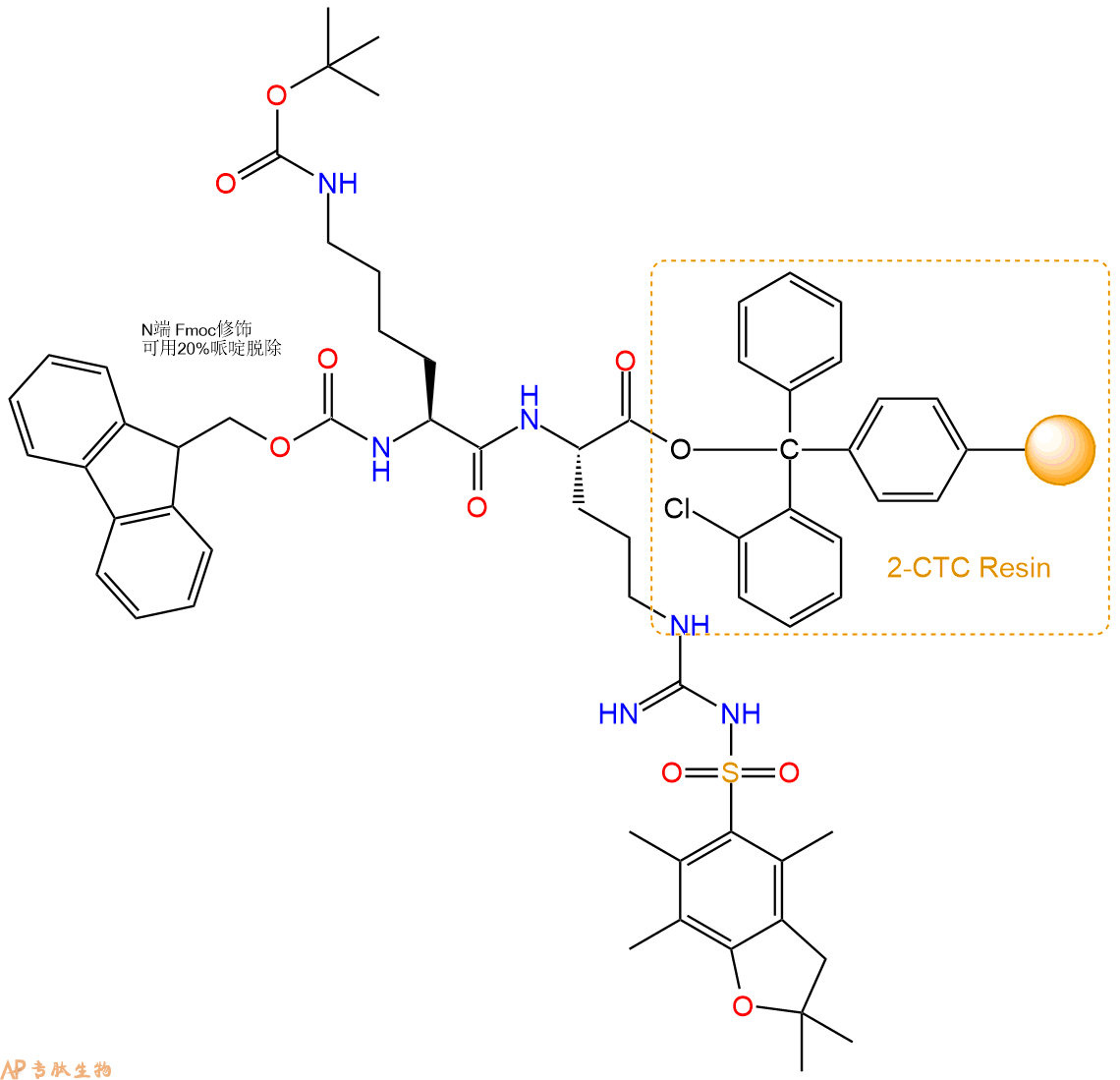
4、依次循环步骤二、步骤三,依次得到
H2N-Lys(Boc)-Arg(Pbf)-CTC Resin
Fmoc-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
H2N-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
Fmoc-Pro-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
H2N-Pro-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
Fmoc-His(Trt)-Pro-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
H2N-His(Trt)-Pro-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
Fmoc-Gln(Trt)-His(Trt)-Pro-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
H2N-Gln(Trt)-His(Trt)-Pro-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
Fmoc-Arg(Pbf)-Gln(Trt)-His(Trt)-Pro-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
H2N-Arg(Pbf)-Gln(Trt)-His(Trt)-Pro-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
Fmoc-Lys(Boc)-Arg(Pbf)-Gln(Trt)-His(Trt)-Pro-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin
以上中间结构,均可在专肽生物多肽计算器-多肽结构计算器中,一键画出。
最后再经过步骤二得到 H2N-Lys(Boc)-Arg(Pbf)-Gln(Trt)-His(Trt)-Pro-Gly-Lys(Boc)-Arg(Pbf)-CTC Resin,结构如下:
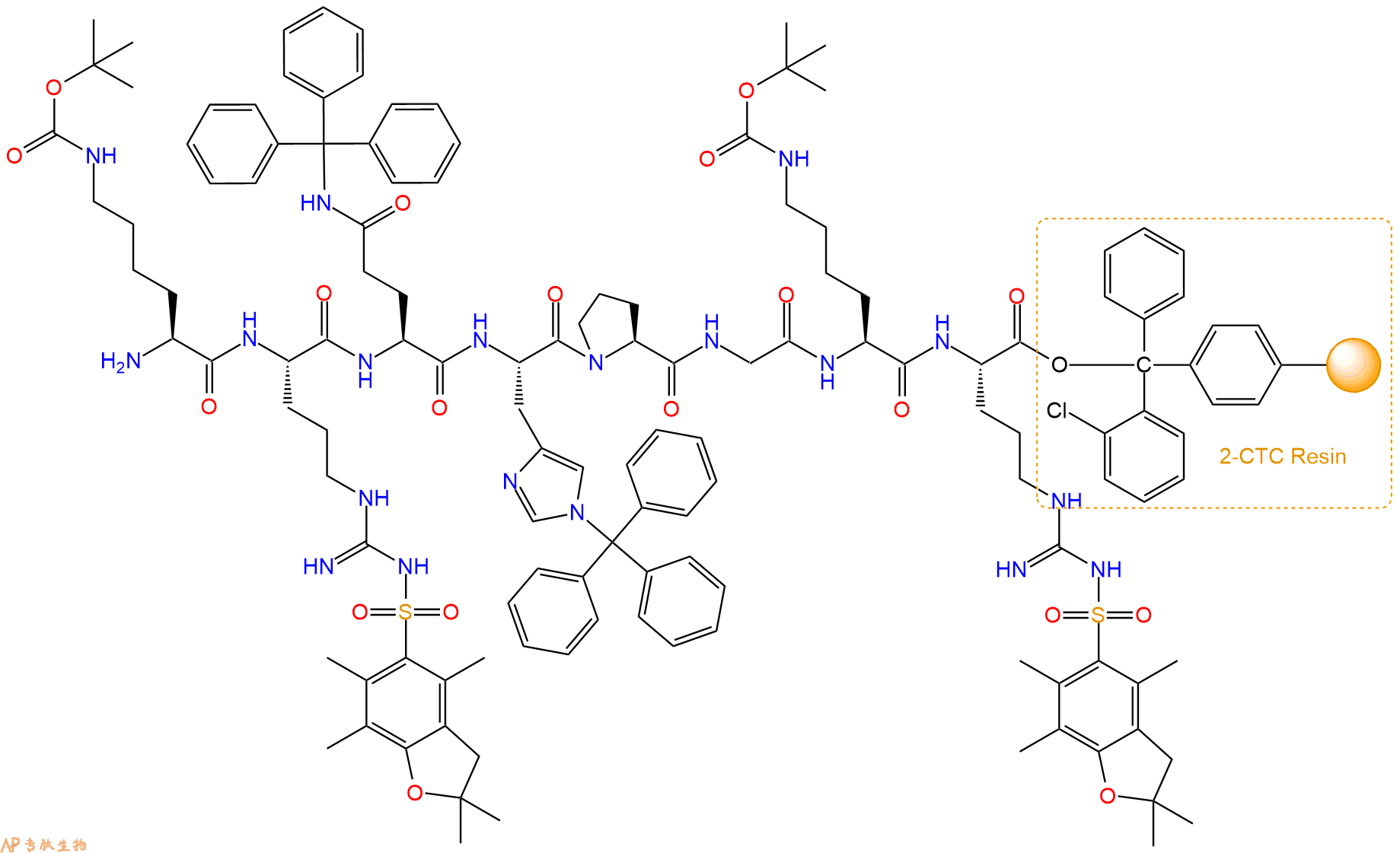
5、切割:6倍树脂体积的切割液(或每1g树脂加8ml左右的切割液),摇床摇晃 2小时,过滤掉树脂,用冰无水乙醚沉淀滤液,并用冰无水乙醚洗涤沉淀物3次,最后将沉淀物放真空干燥釜中,常温干燥24小试,得到粗品H2N-Lys-Arg-Gln-His-Pro-Gly-Lys-Arg-COOH。结构图见产品结构图。
切割液选择:1)TFA:H2O=95%:5%、TFA:H2O=97.5%:2.5%
2)TFA:H2O:TIS=95%:2.5%:2.5%
3)三氟乙酸:茴香硫醚:1,2-乙二硫醇:苯酚:水=87.5%:5%:2.5%:2.5%:2.5%
(前两种适合没有容易氧化的氨基酸,例如Trp、Cys、Met。第三种适合几乎所有的序列。)
6、纯化冻干:使用液相色谱纯化,收集目标峰液体,进行冻干,获得蓬松的粉末状固体多肽。不过这时要取小样复测下纯度 是否目标纯度。
7、最后总结:
杭州专肽生物技术有限公司(ALLPEPTIDE https://www.allpeptide.com)主营定制多肽合成业务,提供各类长肽,短肽,环肽,提供各类修饰肽,如:荧光标记修饰(CY3、CY5、CY5.5、CY7、FAM、FITC、Rhodamine B、TAMRA等),功能基团修饰肽(叠氮、炔基、DBCO、DOTA、NOTA等),同位素标记肽(N15、C13),订书肽(Stapled Peptide),脂肪酸修饰肽(Pal、Myr、Ste),磷酸化修饰肽(P-Ser、P-Thr、P-Tyr),环肽(酰胺键环肽、一对或者多对二硫键环),生物素标记肽,PEG修饰肽,甲基化修饰肽
以上所有内容,为专肽生物原创内容,请勿发布到其他网站上。